Клубни у картофеля появились благодаря гибридизации дикого Solanum etuberosum с томатом

Рис. 1. Картофель (Solanum tuberosum). Общий вид, листья, цветы, клубни. Фото с сайта commons.wikimedia.org
Картофель — привычный продукт, без которого трудно представить современную кухню, — имеет удивительное эволюционное прошлое. Авторы недавно опубликованной в журнале Cell статьи показывают, что культурный картофель и его 107 диких сородичей произошли в результате древнего события гибридного видообразования («скрестились» предки современного томата и южноамериканского растения Solanum etuberosum), а также что само образование клубней — ключевой инновационный признак — имеет гибридное происхождение.
Картофельное пюре, золотистая жареная картошка или тонкие ломтики чипсов — все это разнообразие блюд дает нам один вид растения — культурный картофель (Solanum tuberosum, рис. 1). Сегодня он занимает третье место среди продовольственных культур мира и вместе с пшеницей, рисом и кукурузой обеспечивает большую часть калорий в рационе человека.
Но за привычным клубнем скрывается целый мир. Картофель принадлежит к огромному роду Паслен (Solanum), включающему около 1200 видов, произрастающих от тропиков до умеренных широт. Среди них встречаются и лекарственные растения, и декоративные, и, конечно, важнейшие сельскохозяйственные культуры: картофель, баклажан и томат. Внутри этого обширного рода культурный картофель и его 107 диких сородичей образуют особую эволюционную линию — секцию Petota. Ее ближайшими соседями по эволюционному древу являются две другие линии: Tomato (17 видов, включая культурный томат) и Etuberosum (всего три южноамериканских вида). И Petota, и Etuberosum — геофиты, то есть растения с подземными возобновляющимися органами (рис. 2, А). У Etuberosum это корневища, а у Petota — столоны, от которых отходят клубни. Именно клубни — утолщенные части подземных побегов — сделали картофель столь ценным для человека.
Несмотря на генетическую близость эти три линии хорошо отделены друг от друга: искусственные скрещивания между ними приводят лишь к образованию мелких абортивных семян. К тому же у Petota наблюдается впечатляющее разнообразие уровней плоидности — от диплоидов до полиплоидов, тогда как у его соседей доминируют диплоидные формы. При этом число хромосом (2n = 24) у всех трех линий одинаково, а структура и порядок генов (синтения) удивительно схожи. Установить точные родственные связи между этими линиями долгое время не удавалось. Какая же из них — Etuberosum или Tomato — является ближайшей сестринской группой к картофелю? Данные разных генетических анализов противоречили друг другу, поэтому авторы недавно опубликованной статьи в журнале Cell предприняли масштабное исследование, чтобы разобраться в сложных семейных отношениях в роде Паслен.
Ученые начали с общей картины: построили филогению всего семейства пасленовых (Solanaceae), используя полные геномы 27 видов, охватывающих весь спектр родства — от представителей группы Petota (шесть видов) до Etuberosum, Tomato и еще 19 линий Solanaceae. Казалось бы, такого масштаба данных должно было хватить, чтобы уверенно определить ближайшего «родственника» картофеля. Но результаты оказались парадоксальными: в одних анализах сестринской линией выступал Tomato, в других — Etuberosum. Ни один из вариантов не получил достаточно сильной статистической поддержки (рис. 2, В).

Рис. 2. A — сравнение подземных органов и географическое распространение Tomato (красный), Petota (желтый) и Etuberosum (голубой). Etuberosum формирует корневища. Petota формирует столоны (Stolon) и клубни (Tuber). B — филогенетическое дерево растений рода Паслен (Solanum). Серые линии представляют 500 отдельных филогенетических деревьев, построенных для отдельных участков генома. Линия Petota оказывается сестринской к Tomato (красный, статистическая поддержка 58%) или к Etuberosum (синий, статистическая поддержка 33%). Такая неопределенность в родственных связях может быть свидетельством гибридного происхождения линии Petota. Рисунки из обсуждаемой статьи
Чтобы разобраться в этой путанице, исследователи расширили набор данных: собрали 128 высококачественных геномов, включая 101 геном Petota (44 из 107 известных диплоидных диких видов), 15 геномов Tomato (12 из 16 видов), 9 геномов Etuberosum (все 3 диплоидных вида) и 3 генома внешних групп рода Solanum. Это крупнейший на сегодняшний день набор диких картофельных геномов. Далее все эти геномы выровняли относительно референсного генома картофеля. Результаты оказались показательными: уровень совпадений внутри Petota варьировал от 39,55% до 86,03%, что говорит о высоком уровне генетического разнообразия внутри секции. Для Etuberosum средний уровень составил 32,66%, для Tomato — 23,58%, а для внешних групп всего 11,98%. Однако при построении эволюционных деревьев исследователей ждал новый сюрприз: разные участки ДНК рассказывали разные истории. Одни гены уверенно указывали Tomato как ближайшего родственника Petota, другие — Etuberosum, а третьи и вовсе показывали, что Tomato и Etuberosum — сестринские линии, а Petota отстоит чуть дальше.
Такой беспорядок в свидетельствах мог возникнуть по двум причинам: либо из-за случайного наследования генов от далекого общего предка (неполная сортировка линий, см. Incomplete lineage sorting), либо из-за древнего скрещивания между уже разделившимися видами (гибридизация).
Чтобы проверить, какой вариант ближе к истине, исследователи разбили геномы на тысячи небольших фрагментов по 100 тысяч пар оснований и для каждого построили свое мини-«древо». Всего получилось 3436 таких деревьев, которые в основном показывали три характерных варианта родословных. Половина из них показывала наиболее близкое родство Petota с Tomato (Tree-I), 37% — с Etuberosum (Tree-II), а около 12% поддерживали родство Tomato и Etuberosum между собой (Tree-III). Если бы имела место только неполная сортировка линий, то три альтернативных дерева (Tree-I, Tree-II, Tree-III) должны были бы встречаться в геноме в примерно одинаковых пропорциях, так как случайное наследование разных аллелей от общего предка не должно иметь предпочтения к какой-то топологии. Но заметное преобладание первых двух топологий над третьей указывает на гибридное происхождение. Также исследователи проанализировали не случайные участки ДНК, а отдельные гены (более 17 тысяч генетических семейств). Деревья были построены по кодирующим последовательностям, белковым последовательностям, а также по экзонным и интронным областям. И снова получилась та же картина: три основных варианта деревьев, причем деревья I и II встречались значительно чаще по сравнению с деревом III (рис. 3).

Рис. 3. Сравнение разных филогенетических сценариев. Слева схематически показаны три возможные топологии (варианта расположения ветвей), отражающие родственные отношения между тремя группами (Petota, Tomato, Etuberosum). Справа приведены столбчатые диаграммы с подсчетом, какая топология (Tree-I — красный, Tree-II — зеленый, Tree-III — желтый) чаще встречается при анализе генов (Gene), кодирующих последовательностей (CDS), белков (Protein) и интронов (Intron). Оказалось, что топология Tree-II встречается значительно чаще, чем Tree-III. Рисунок из обсуждаемой статьи
Затем авторы провели модельные симуляции в виртуальных популяциях, учитывая только неполную сортировку линий. Когда они сравнили результаты этих симуляций с реальными данными, разница оказалась слишком велика: реальные геномы содержали больше сигналов смешения, чем могла объяснить простая сортировка древнего разнообразия. Более того, сравнив закономерности распределения предковых и производных аллелей (см. ABBA-BABA test) авторы обнаружили следы древнего потока генов между между картофелем (Petota) и различными образцами Tomato и Etuberosum. Это прямое доказательство того, что гены перемещались между этими группами уже после того, как их эволюционные пути разошлись.
Следующий вопрос, который необходимо было прояснить: идет ли речь о единичном древнем событии, из которого вышла вся линия Petota, или же о множественных интрогрессиях, затрагивавших лишь отдельные виды? Для этого авторы проанализировали 349 геномов — диких и культурных форм картофеля. Оказалось, что во всех случаях доля «наследства» от Etuberosum составляет примерно 60%, а от Tomato — около 40%. Такое соотношение сохраняется у всех клад Petota, что говорит не о частных эпизодах скрещивания, а о едином древнем событии гибридизации, предшествовавшем разделению картофельной линии на виды. Когда исследователи сравнили конкретные участки генома, оказалось, что совпадения с Etuberosum встречаются у всех трех основных клад Petota в одних и тех же местах. Если бы скрещивания происходили неоднократно и в разное время, у разных видов эти участки были бы разными.
Если линия Petota действительно возникла в результате гибридизации, то время ее расхождения с каждой из родительских ветвей должно быть почти одинаковым и, разумеется, более молодым, чем время разделения самих родительских линий. Оценка времени расхождения между линиями показала, что Petota расходится с Tomato примерно 9 миллионов лет назад, а с Etuberosum — около 8,5 миллиона лет назад. Эти даты практически совпадают друг с другом, но при этом существенно моложе расхождения Tomato и Etuberosum (около 13 миллионов лет назад). Это отлично вписывается в сценарий гибридного происхождения. Компьютерное моделирование, учитывающее как разделение линий, так и возможный обмен генами не только подтвердило время гибридизации (8,6 млн лет назад), но и позволило также количественно измерить доли вклада родительских линий: примерно 51% генома пришел от предка Tomato, а 49% — от предка Etuberosum (рис. 4). Это событие совпало по времени с бурным поднятием Анд (6–10 млн лет назад). Вероятно, масштабные геологические изменения создали экологические ниши, где гибридное происхождение стало ключевым преимуществом, позволившим картофелю освоить новые местообитания.
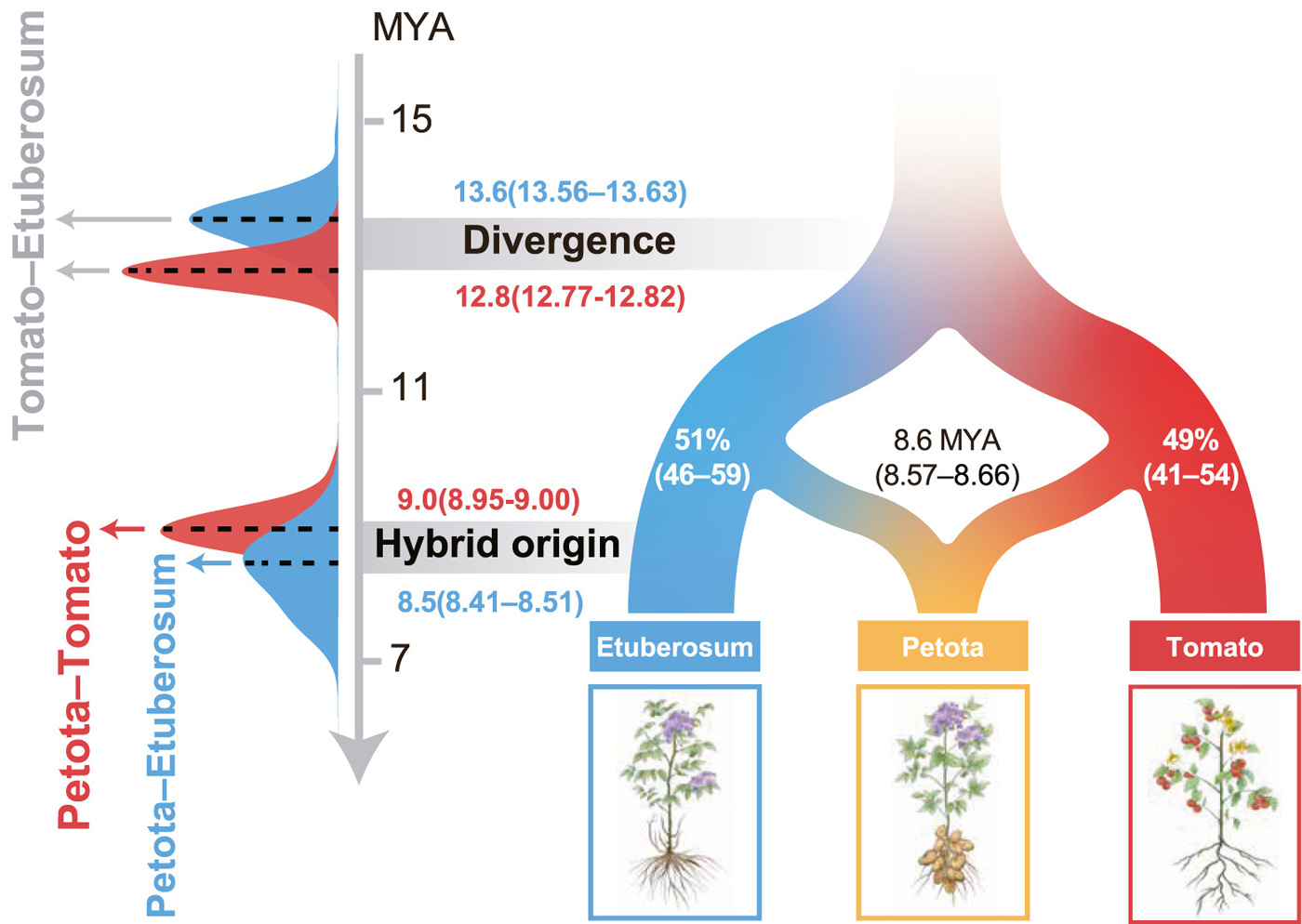
Рис. 4. Схематическое изображение эволюционной истории рода Petota: около 13 млн лет назад произошла дивергенция (Divergence) линий Etuberosum и Tomato; около 8,6 млн лет назад произошла гибридизация (Hybrid origin), в результате чего возникла линия Petota, ставшая основой для всех современных диких картофелей. Вклад родительских линий в формирование гибрида составляет 51% от Etuberosum и 49% от Tomato. Рисунок из обсуждаемой статьи
И Petota, и Etuberosum образуют подземные побеги — столоны или корневища. Но только у Petota концы столонов утолщаются и превращаются в клубни — запасающие органы, которые мы привыкли видеть на столе в виде картофелин. Исследователи предположили, что это эволюционное новшество стало возможным благодаря гибридизации. Когда в одном геноме встретились гены двух разных линий Tomato и Etuberosum, возникла новая сеть взаимодействий, которая и открыла путь к созданию принципиально нового органа. Чтобы проверить эту гипотезу, ученые проанализировали тысячи генов и обнаружили: ключевые элементы клубнеобразования имеют мозаичное происхождение. Одни пришли от Tomato, другие — от Etuberosum, и вместе они сложились в единую систему. В итоге получилась сеть, где гены разного происхождения стали ключевыми деталями для сборки клубня.
Затем авторы перешли к экспериментам и сосредоточились на двух генах, связанных с клубнеобразованием: SP6A (от Tomato) и IT1 (от Etuberosum). Этот комплекс запускает образование клубня. Чтобы проверить, могут ли белки разных линий взаимодействовать, ученые сначала применили дрожжевой двугибридный анализ. Если белки образовывали комплекс, то за счет слияния сшитых с ними транскрипционных доменов происходило включение гена-репортера, и дрожжи успешно росли на среде без необходимых аминокислот. В клетки внедряли разные версии генов SP6A и IT1 из Petota, Tomato и Etuberosum и смотрели, какие комбинации дают результат. Помимо этого, применили метод «светящихся белков». В листья табака (Nicotiana benthamiana) внедряли те же самые белки, сшитые с половинками фермента люциферазы (того самого, что позволяет светлячкам светиться). Если белки образовывали комплекс, половинки фермента соединялись, и в листьях появлялось свечение, видимое через специальную камеру. Таким образом удалось буквально увидеть взаимодействие белков в живой клетке.
Оба эксперимента однозначно показали, что белок SP6A из Petota способен образовывать комплекс с IT1. У Etuberosum же SP6A «сломался»: он не связывался с IT1 и к тому же нигде не экспрессировался. Зато IT1 у Etuberosum оставался рабочим и включался в корневищах, но без партнера в лице SP6A клубни не формировались. Интересно, что IT1 из картофеля и Etuberosum действовали практически одинаково, как полноценные «молекулярные выключатели», а вот версия IT1 у Tomato была куда менее активной. Получается, что картофель унаследовал «переключатель» IT1 от Etuberosum и полноценную «сигнальную молекулу» SP6A от Tomato, и именно эта комбинация дала старт механизму образования клубней.
Дальше исследователи проверили и другие гены — DRN и CLF, в которых нашли следы гибридного происхождения. У Petota в DRN оказалось 23 мутации (9 от Etuberosum и 14 от Tomato), а в CLF — тоже 23, но уже с другим соотношением (17 от Etuberosum и 6 от Tomato). При выключении у растений дикого картофеля Solanum chacoense с помощью CRISPR-Cas9 (см. картинку дня CRISPR/Cas-системы) гена DRN формировались лишь крошечные недоразвитые клубни, а при нокауте гена CLF растения становились карликовыми и вовсе не образовывали подземных органов (рис. 5). Таким образом, гибридизация не просто «смешала» геномы Tomato и Etuberosum, а предоставила обширный запас генетических вариаций, из которых естественный отбор собрал оптимальные комбинации. В итоге появился новый орган — клубень. Вегетативное размножение с помощью клубней не только закрепило полезные генетические сочетания, но и, вероятно, усилило эффект гетерозиса — то есть повышенной жизнеспособности гибридов.

Рис. 5. Фенотипы растений дикого картофеля Solanum chacoense дикого типа (WT) и мутантов с выключенными генами DRN (слева) и CLF (справа). Рисунок из обсуждаемой статьи
Все дикие родственники картофеля объединяются в группу Petota, и у всех 107 видов этой линии есть клубни. При этом Petota показывает самую быструю скорость видообразования в своей группе. По расчетам авторов, новые линии появлялись здесь со скоростью 0,53 вида на миллион лет. Для сравнения: у Etuberosum этот показатель равен 0,39, у Tomato — 0,44, а в среднем по роду Solanum — всего 0,38. Получается, что именно линия картофеля «разбежалась» по эволюционному древу гораздо быстрее других. Ученые предположили, что толчком к этому стала именно древняя гибридизация. Когда гены двух разных предковых линий оказались объединены в одном геноме, возник необычайно богатый генетический багаж. Такой запас вариаций позволил появляться новым комбинациям признаков и дал гибридным видам возможность «перебирать» разные решения в ответ на изменения среды и осваивать новые экологические ниши, возникшие в результате подъема Анд.
Авторы проанализировали 4703 точечных наблюдения для трех видов Etuberosum, всех 16 диких видов Tomato и всех 107 некультивируемых видов Petota и установили, что дикие виды Petota встречаются в более широком диапазоне условий и занимают гораздо разнообразные экосистемы — от высокогорных альпийских лугов в Андах до сезонно-сухих тропических лесов. Наибольшая их концентрация наблюдается в холодных горах Центральной Мексики и в центральных Андах. Если сравнивать климатические предпочтения, то ареалы Petota в среднем суше, чем у Etuberosum, но холоднее и более влажные, чем у диких Tomato. По разнообразию экологических ниш картофельные виды тоже впереди: их «экологическая широта» более чем вдвое превышает показатели Tomato и почти в четыре раза — Etuberosum.
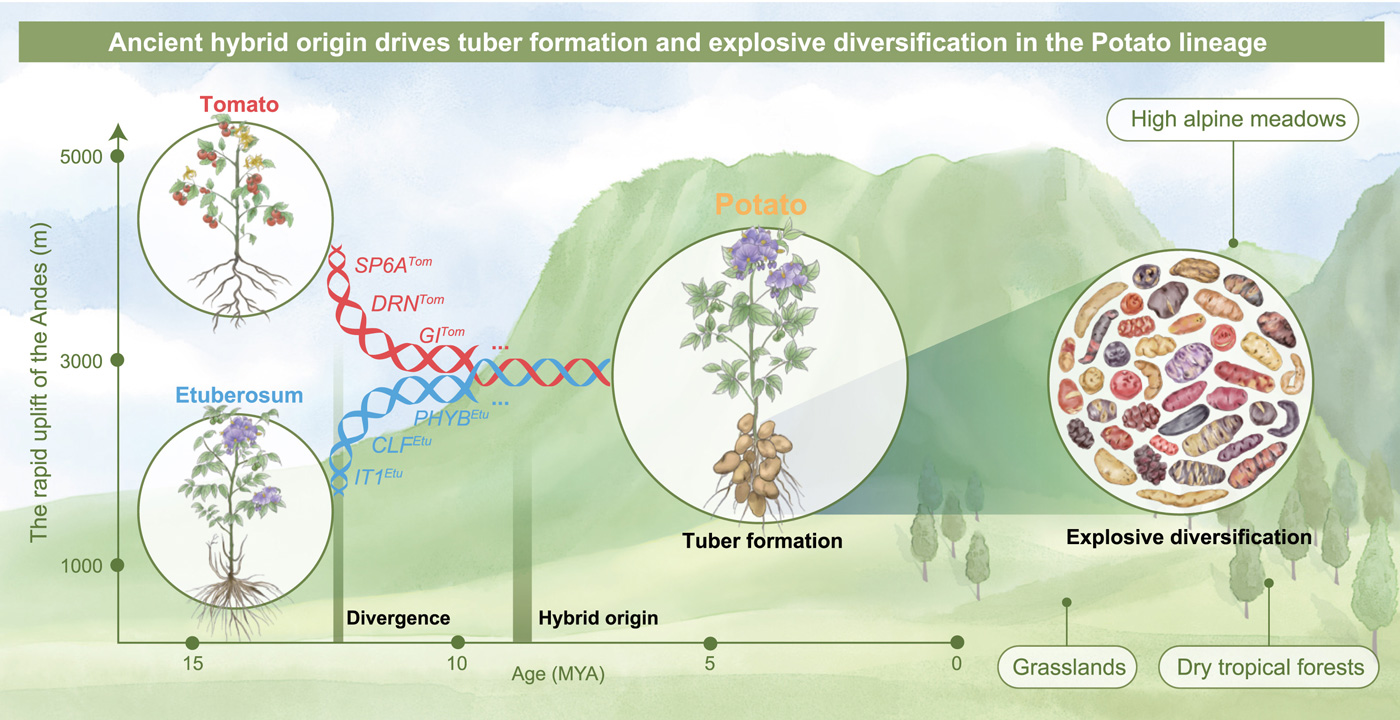
Рис. 6. Эволюционная история картофеля. Картофель (Potato) возник в результате древней гибридизации линий Tomato и Etuberosum (Hybrid origin), произошедшей около 8–9 млн лет назад, что совпало с быстрым подъемом Анд (6–10 млн лет назад). Цветные стрелки с генами (SP6A, DRN, GI, PHYB, CLF, IT1) показывают, какие гены пришли от предков томата (красные), а какие — от предков линии Etuberosum (синие). Гибридное происхождение обеспечило формирование клубня (Tuber formation) и дало богатый набор генов, что привело к «взрывной» диверсификации (Explosive diversification) и освоению самых разных местообитаний. По вертикальной оси — высота над уровнем моря (до 5000 м), отражающая разнообразие местообитаний: от тропических лесов (Dry tropical forests) и лугов (Grasslands) до высокогорных альпийских лугов Анд (High alpine meadows). По горизонтальной оси — время в миллионах лет (MYA). Рисунок из обсуждаемой статьи
В итоге складывается цельная история (рис. 6). Гибридизация между Tomato и Etuberosum создала широкий спектр генетических вариаций. Это обеспечило возможность формирования клубня — нового органа, возникшего в результате объединения генетических элементов обоих родительских линий. Возникновение клубня стало ключевым фактором эволюционного успеха картофеля. Сочетание новой морфологии с гибридным геномом и давлением естественного отбора привело к быстрой диверсификации видов и позволило диким видам картофеля освоить широчайший спектр условий — от тропических лесов до суровых высокогорий Анд. И, пожалуй, зная всю эту эпичную сагу о гибридах, эволюционной радиации и клубнях, самое логичное завершение — отправиться на кухню и пожарить картошечку, теперь уже с полным осознанием ее невероятного прошлого.
Источник: Zhiyang Zhang, Pingxian Zhang, Yiyuan Ding, Zefu Wang, Zhaoxu Ma, Edeline Gagnon, Yuxin Jia, Lin Cheng, Zhigui Bao, Zinan Liu, Yaoyao Wu, Yong Hu, Qun Lian, Weichao Lin, Nan Wang, Keyi Ye, Hongru Wang, Jinzhe Zhang, Yongfeng Zhou, Liang Liu, Suhua Li, William J. Lucas, Tiina Särkinen, Sandra Knapp, Loren H. Rieseberg, Jianquan Liu, Sanwen Huang. Ancient hybridization underlies tuberization and radiation of the potato lineage // Cell. 2025. DOI: 10.1016/j.cell.2025.06.034.
Елена Устинова